
DRUGAI
今天为大家介绍的是来自Ken A. Dill团队的一篇论文。生命起源时,特定有用的蛋白质序列是如何从更简单的分子中产生的?这个看似“在大海捞针”的问题,与早期的“蛋白质折叠问题”有着惊人的相似,而后者的解决方案现在已经从统计物理学中得出。基于这样一个逻辑:生命的起源一定是在有一种作用于表型而非基因型的演化机制出现之后,作者认为蛋白质及其折叠过程很可能是生命起源早期阶段的主要推动因素。

目前没有人知道生命是如何起源的——大约在35亿到38亿年前的地球上产生。迄今为止,没有任何实验能够重现这一过程。这个问题可以进一步聚焦在核酸和蛋白质上,因为这两者是生物学核心中的两种具有“序列→结构→功能”特性的分子。一种流行的观点认为RNA首先出现,因为它既能储存信息又能催化反应,从而实现自我复制。在这里,作者总结了一个近期的不同观点,认为蛋白质的折叠和功能可能是生命起源的最初步骤之一。
为什么是蛋白质,而不是RNA?认为RNA优先的观点基于“假设自我复制是核心问题”这一假设。而认为蛋白质优先的观点则基于另一种推理:生命的驱动力究竟是什么?为什么会有这样的驱动力存在?哪种分子过程是从无序状态开始的,并通过某种“大海捞针”式的搜索在巨大的序列空间中找到少数几个具有功能的生物分子序列?作者描述了生命起源问题中的“大海捞针”性质,与解决另一个看似同样复杂的“大海捞针”难题——蛋白质折叠问题所涉及的基本物理过程非常相似,并且可以用这一物理过程来解释。
生命起源的基本问题
进化是如何开始的?根据NASA的定义,“生命是能够自我维持并具备达尔文进化能力的化学系统”。作者在此特别强调这一定义中的隐含意义:因为生命无法在缺乏适应性动态(adaptation dynamics)的情况下被定义,那么这种动态形式必定在生命起源时或更早已经存在。简而言之,生命只有在能够传播的情况下才能起源。进化必须有一个开端。显然,在大约35亿年前生命起源之前,分子之间的达尔文过程并未发生,那时地球的过程完全由物理和化学规律支配。在生命系统能够挑选分子之前,必定存在一种可持续的筛选过程。那么,这种过程是什么呢?以下是一些关键问题。
1、聚合物序列是如何开始编码分子功能的?生物学的核心在于基于序列的异聚物(RNA、DNA、蛋白质),它们是细胞的催化剂、机器和记忆。生命的起源有时被比喻为在大海捞针,或像“盲眼钟表匠”,或像猴子在打字机上敲出莎士比亚的剧本,因为赋予生物聚合物功能的特定序列必须从绝大多数无用的替代序列中找到。聚合物序列中的有序是如何从无序中产生的?
2、从降解和水解到长期稳定的转折点是什么?原始生命化学反应通常倾向于水解、降解和稀释,这符合热力学第二定律的平衡原则。然而,生命系统并不追求平衡状态,而是通过资源摄入来维持自身。那么,原始分子是如何发展出克服衰变力量的持续动态的?
3、在细胞出现之前,适应性是什么?生物学通过在有限资源的竞争中胜出或失败来推动自我利益的实现。然而,简单的原始分子并没有显而易见的自我利益表现。在细胞中观察到的适应性景观攀升之前,分子之间遵循的是什么样的选择原则?
蛋白质的大海捞针
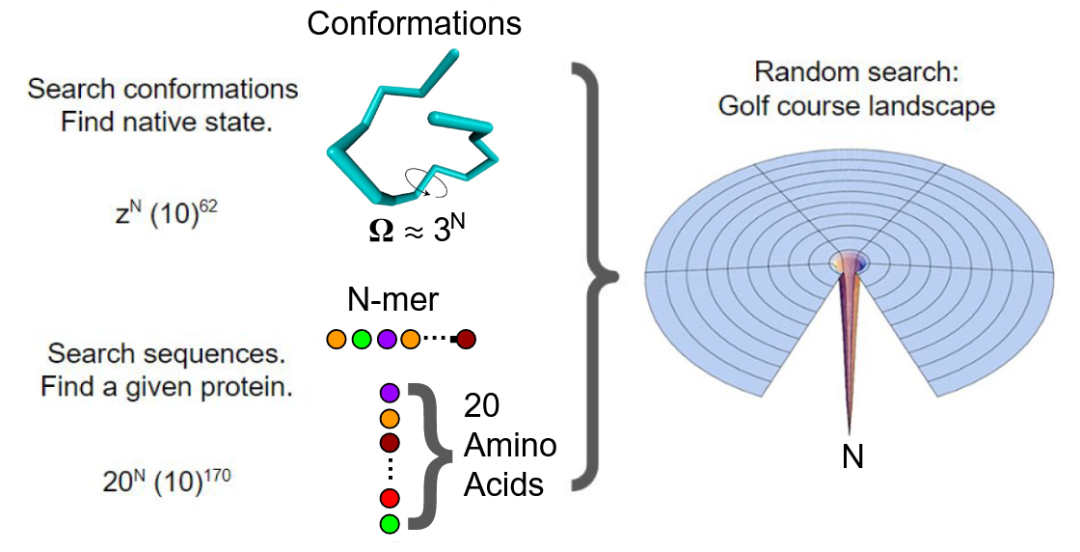
图 1
蛋白质折叠问题(PFP)以前也被视为“大海捞针”的搜索问题(图1左上):蛋白质分子如何在其巨大的构象空间中找到其唯一的本征结构(native structure)?这个问题也可以用高尔夫球场形状的景观来表达。然而,驱动力和动力学路径的物理折叠问题的研究表明 “大海捞针”和高尔夫球场的比喻现在被视为不正确的概念化。问题并不在于随机的独立步骤,关键在于找到哪种类型的物理协同作用能够导致一个小概率状态向更高概率状态的雪球效应或自我增强。
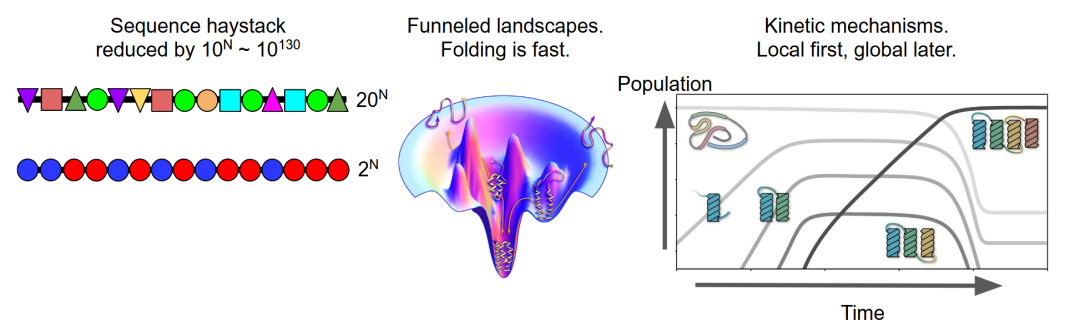
图 2
图2展示了蛋白质折叠问题的三个启示:1)基于疏水(H)和极性(P)模式的物理编码减少了“大海捞针”的难度,超过100个数量级,这已被实验验证。2)由于二级和三级驱动力中的物理协同性,折叠景观相对呈漏斗状,而不是像高尔夫球场。3)动力学是先局部后整体的,先形成螺旋和转角,后形成三级结构,这将NP-completeness挑战简化为一个非常快速的过程。在Foldon漏斗模型中,早期快速步骤(螺旋和转角)并不稳定,它们只是相对不那么不稳定,沿着一个逐渐平缓的景观继续上升,直到达到一个临界点,在该点上本征结构实现了完全稳定。要通过随机搜索所有个长度为N的序列空间来找到特定蛋白质的序列(如溶菌酶),概率为。但由于二元HP折叠编码,搜索特定折叠和功能的空间只需搜索个序列(见图2左侧);这个空间缩小了一个的数量级,非常显著的缩小。
假设:进化始于蛋白质
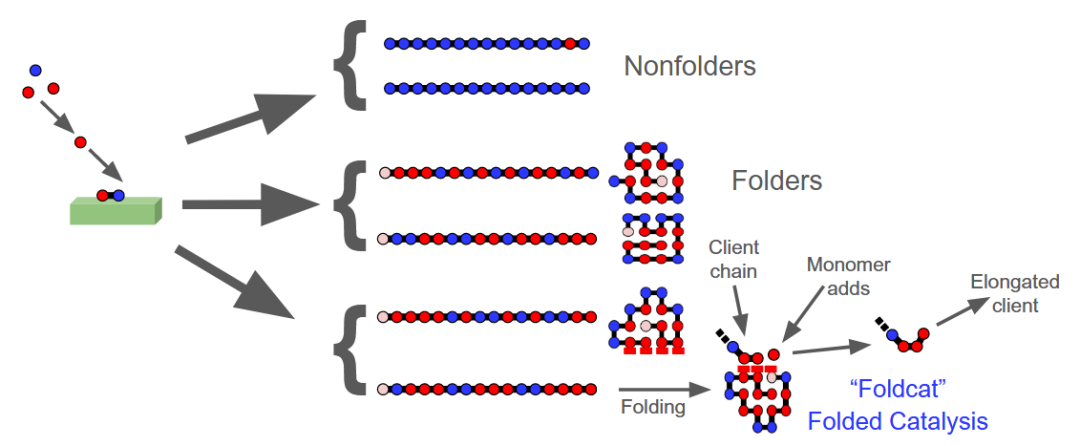
图 3
HP Foldcat 机制。现在,讨论前生物进化与蛋白质折叠过程之间的联系,这一过程被称为 HP Foldcat(HPF)机制。HP Foldcat 机制的详细介绍见其他文献,这里做一个总结。“HP”代表疏水性(hydrophobic)和极性(polar),这两种类型的氨基酸单体可以聚合成链。假设在某种生命起源前的催化剂上(称为Founding Rock;见图3)通过催化作用,这些单体不断合成肽。起初,这些肽链是短的且随机的。其中有一小部分较长的肽链会折叠成紧密的构象,形成一个疏水核心。有些折叠体具有稳定的表面,被称为具有疏水性“着陆垫”的foldcat序列,这些序列是催化剂,通过将下一个单体推进链中,加速客户链(client chain)的延长。以下部分首先是模型的整体结论,然后是对其更量化的描述。
Foldcat过程具有涌现(Emergent)特性,即,这些特性不是单纯的、非协同的随机短链合成所能预期的。a)链条变得更长。b)存在自催化集合,其中一些序列优先积累,这解释了序列与结构关系的起源。c)出现了一种适应性特性,即折叠稳定性(最终是催化效率),通过这种特性,一些序列得以生存和胜出,而其他序列则降解和循环再生。尽管适应性最初表现为简单的折叠稳定性,但一旦这种优势形式得到持续的巩固,任何其他能够稳定蛋白质或其群体的因素也能支持进一步的进化变化。d)相应地,一个类似进化的过程出现了,即通过单体竞争进行的序列搜索和适应性选择;自催化和功能的涌现在这里不依赖于核酸模板。e)此外,该机制具有长期持续性,原因如下:1)它是一个开放系统,不受第二定律趋向平衡的驱动,而是由单体的不平衡输入和founding rock驱动的,founding rock最初促进了这些单体的非自发聚合。2)在过程的某个阶段,当链条变成足够好的foldcat时,将发生“解缆”( untethering)转变,即在这个时间点,催化主要由foldcat蛋白进行,不再依赖于founding rock。这将是一个重大的进化事件,因为起源不再是局限的,例如,不再“困于内布拉斯加州的小池塘”里。现在,催化剂是可移动的,可以去任何地方;现在,催化剂是可编程的(不同的蛋白质可以催化不同的反应或在不同条件下工作);现在,催化作用被微型化,并且可以在细胞内捕获。这些涌现特性来源于两种物理协同作用:i)足够长的链条具有折叠的核心,这些核心对降解有更强的抵抗作用;ii)帮助其他链条延长的链条集合形成了自催化集合。就像雪花不断积累成雪球,最终变成雪崩一样。协同性是解释“微小的第一步如何逐渐占据宏观尺度”的关键。HP Foldcat 机制就像雪球一样,逐渐发展出更长、更折叠、催化性更强的分子。
该机制在生物起源前具有合理性。虽然未被直接观察到,但HP Foldcat机制很可能在早期地球上出现。首先,氨基酸和肽已经通过地球上的过程或来自太空的方式以生物起源前合理的方式生成过。类似founding rock的催化作用,即通过脱水反应延长肽链,已在矿物表面和空气-水界面(如海洋飞沫)上得到证明,甚至可能通过未知的地外过程实现。肽键形成的合理非平衡驱动因素可能包括湿干循环或冷热循环。其次,疏水性和极性单体的聚合物可以折叠并催化,即使这些链是随机的或短的。众所周知,蛋白质是由二进制HP码驱动的。由于今天的20种氨基酸在蛋白质数据银行(PDB)中大致以相等的疏水性和极性比例存在,这意味着任何足够长度的序列大多数都会在水中折叠成紧凑的结构,从而其核心部分能够避免与外部溶剂接触。虽然有推测认为早期的氨基酸“字母表”可能少于20种,但这里重要的是仅仅有二进制代码。此外,短蛋白质在生物学中普遍存在:人类体内存在数千种微蛋白,这些微蛋白的长度小于100个氨基酸。虽然现代微蛋白可能有着不同且更晚的进化起源,但它们证明了短蛋白可以满足HP Foldcat机制的需求而具有有趣且功能性的特性。许多微蛋白执行各种生物功能,包括催化反应。第三,折叠和催化是HP链条的简单物理特性。这些特性在氨基酸的生物起源前的混合物中存在,并且可能得到某些小分子的协助。此外,即使只有半胱氨酸这种单一的疏水性氨基酸,也被认为在合理的生物起源前的条件下是可获得的,并能够进行肽连接反应。另外,在淀粉样蛋白中也观察到了催化作用。尽管今天的酶催化剂通常利用高度精细的化学和空间特异性,但正如这里设想的,更简单的空间邻近效应就能够使反应速度提高数个数量级。HP Foldcat机制预测,HP链条的持续生成可能会导致一些较长的折叠链条,其中一部分能够催化其他反应。
编译 | 黄海涛
审稿 | 曾全晨
参考资料
Kocher, C. D., & Dill, K. A. (2024). Origins of Life: The Protein Folding Problem all over again?. Proceedings of the National Academy of Sciences, 121(34), e2315000121.
内容中包含的图片若涉及版权问题,请及时与我们联系删除

评论
沙发等你来抢