
DRUGONE
脂质在膜双层两侧之间的自发跨膜转移(flip-flop)是维持生物膜不对称性的关键过程,但其发生频率极低,实验和传统分子动力学方法均难以直接捕捉其分子机制。研究人员引入一种由人工智能引导的跃迁路径采样方法(AIMMD),在不施加任何偏置力或预设反应坐标的情况下,高效采样脂质翻转与膜纳米孔形成的稀有事件。结果揭示了多种脂质翻转机制,包括干燥隧穿、水纳米孔介导、水线与水纳米液滴辅助等路径,并表明在高维特征空间中,反应坐标可被有效简化为近似线性的形式,为理解膜相关稀有事件提供了新的计算框架。

生物膜通常具有显著的脂质组分不对称性,这一特性对于细胞信号传导、膜蛋白功能和细胞命运调控至关重要。维持这种不对称性需要主动转运机制,而在缺乏相关蛋白的情况下,自发脂质翻转往往极其缓慢,其时间尺度可达分钟级。
脂质翻转的主要能垒来自极性头基穿越疏水膜核心所需的高能量代价。以往研究提出了多种可能机制,包括脂质单独穿越膜核心的“干燥隧穿”路径,以及借助瞬时水孔的“湿润通道”路径。然而,由于事件稀有、反应坐标复杂,传统增强采样方法往往依赖人为设定,难以无偏地识别真实主导机制。
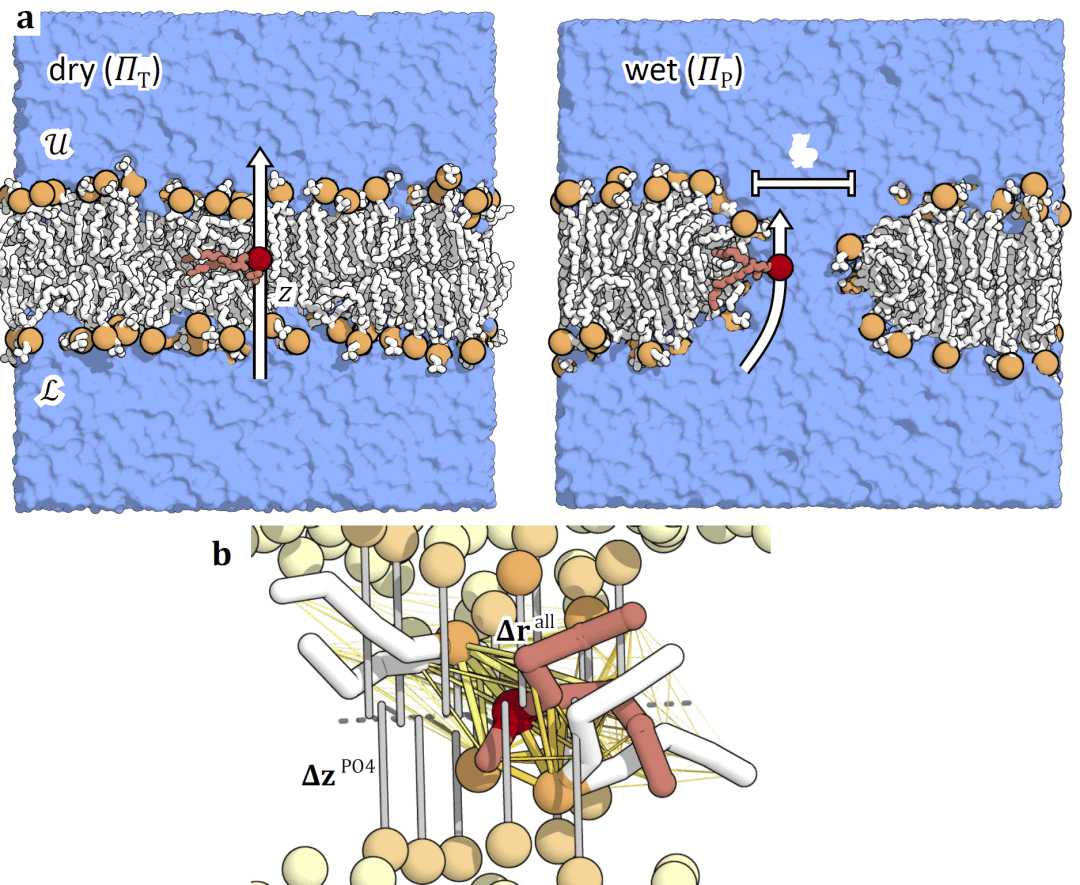
图 1| 脂质翻转(flip-flop)分子动力学模拟。
方法
研究人员采用 AIMMD 框架,将跃迁路径采样与深度神经网络相结合,在分子动力学模拟过程中在线学习脂质跨膜转移的“承诺概率”。模型以脂质位置、取向、邻近脂质排布及局部水化状态等高维特征为输入,引导采样集中于反应过渡区域,从而在全原子与粗粒化模型中高效收集大量脂质翻转与膜孔形成的反应轨迹,而无需预先指定反应路径或施加外部偏置。
结果
粗粒化模型中脂质偏好“干燥隧穿”翻转
在 Martini 粗粒化 DMPC 膜模型中,跃迁路径采样结果显示,脂质主要通过不伴随水孔形成的干燥隧穿机制完成翻转。神经网络分析表明,脂质与邻近脂质之间的相对几何关系比单纯的垂直位移更能准确描述该过程。
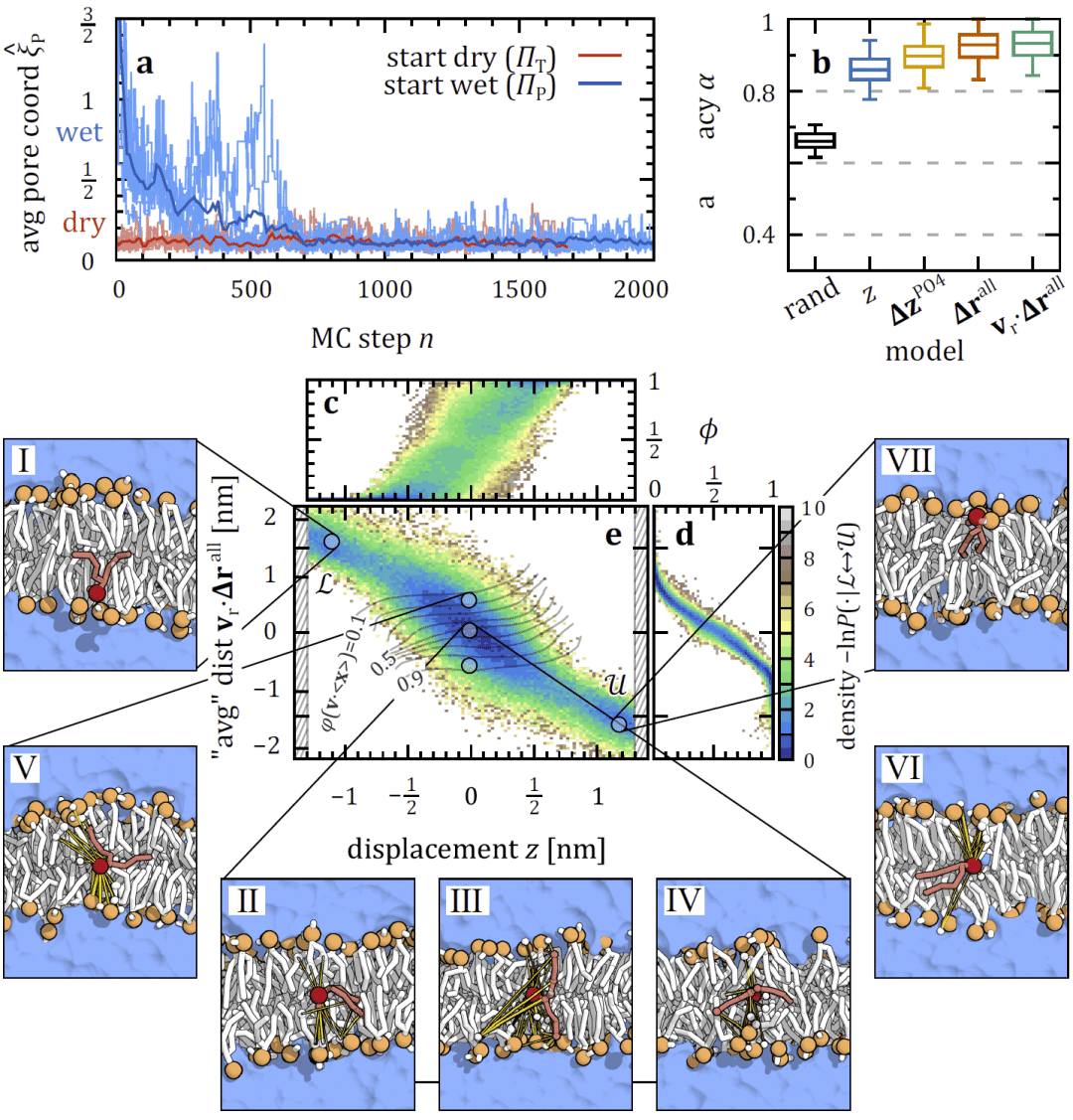
图 2| 粗粒化 DMPC 脂质通过膜内“隧穿”机制完成翻转。
全原子模型中脂质翻转伴随水纳米孔形成
在全原子 DMPC 膜模型中,研究人员观察到脂质翻转几乎总是与自发形成的水纳米孔相关。纳米孔一旦形成,通常可持续足够长的时间,使多个脂质完成跨膜转移,表明该路径在原子级模型中占主导地位。
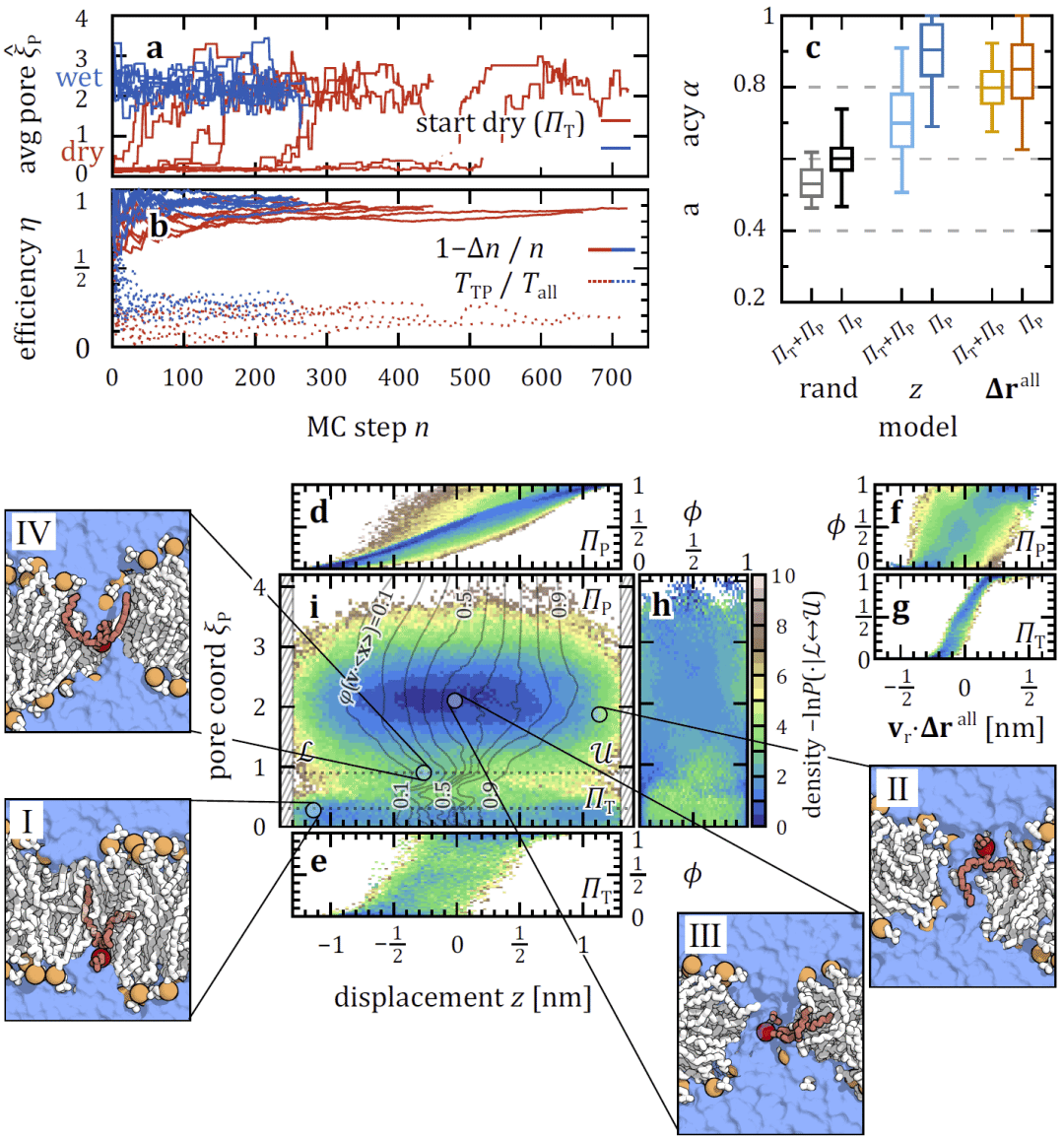
图 3| 全原子 DMPC 脂质经由充水纳米孔实现跨膜翻转。
膜孔形成先于脂质翻转发生
通过专门采样膜孔成核过程,研究人员发现水孔的形成通常先于脂质翻转事件。只有在孔结构维持足够时间的情况下,脂质才会偶然通过孔道完成跨膜迁移,说明膜孔是翻转的必要但非充分条件。
厚膜体系中的“水线 + 局部变薄”机制
在由长链 DSPC 脂质构成的较厚膜中,研究人员识别出一种介于干燥隧穿与完整水孔之间的中间机制。脂质翻转伴随局部膜变薄,并通过短暂形成的水线或水纳米液滴完成头基跨越。
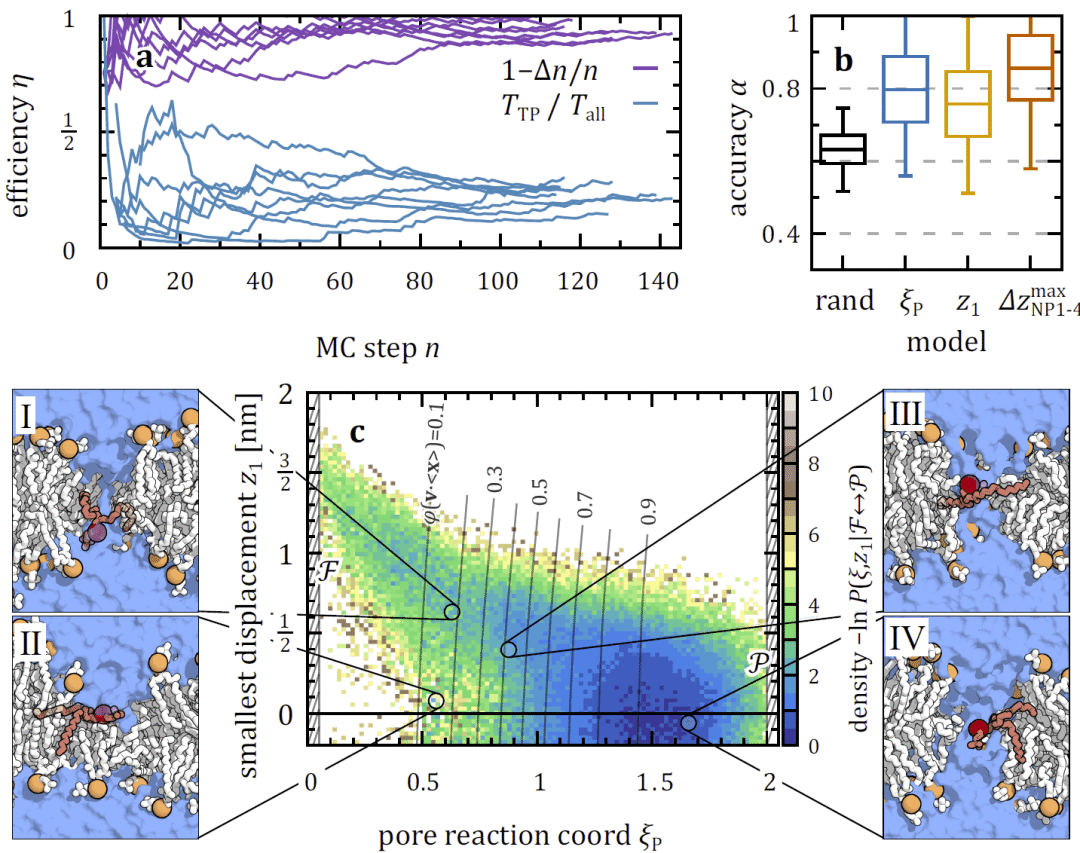
图 4| 基于 TPS 的 Charmm36 DMPC 膜中水纳米孔成核过程采样。
真实质膜中脂质类型决定翻转路径
在哺乳动物质膜模拟体系中,疏水性较强的胆固醇主要采用干燥隧穿机制,而具有极性头基的磷脂则更倾向于借助水线或水液滴完成翻转,显示脂质化学性质对机制选择具有决定性影响。
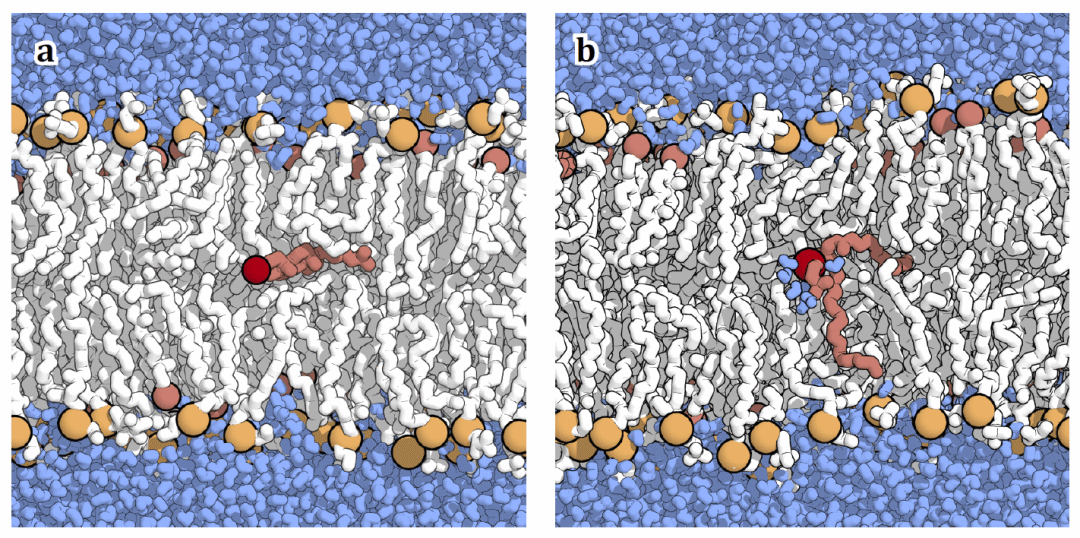
图 5| 全原子分辨率下质膜模拟体系中脂质翻转的过渡态(𝜙≈0.5)。
讨论
该研究系统揭示了脂质翻转并非单一机制,而是在不同膜厚度、脂质类型和模型分辨率下呈现多种竞争路径。AIMMD 方法不仅能够无偏地捕捉这些稀有事件,还在高维特征空间中发现反应坐标呈现近似线性结构,这一现象与高维空间中线性可分性的理论预期相一致。
研究人员提出,生物膜中自发脂质翻转更可能通过短暂的水线或局部极性通道完成,而非稳定水孔或完全干燥的隧穿路径。该工作为理解膜稳定性、电穿孔、脂质不对称性维持及膜蛋白辅助转运机制提供了重要的物理图景,也展示了人工智能在揭示复杂分子机制方面的独特优势。
整理 | DrugOne团队
参考资料
Post, M., Hummer, G. AI-guided transition path sampling of lipid flip-flop and membrane nanoporation. Nat Commun (2025).
https://doi.org/10.1038/s41467-025-67599-3

内容为【DrugOne】公众号原创|转载请注明来源
内容中包含的图片若涉及版权问题,请及时与我们联系删除

评论
沙发等你来抢